Nous avons généré des données de séquençage à l'échelle du génome pour 102 chasseurs-cueilleurs nouvellement signalés et augmenté la couverture pour 14 individus précédemment publiés 4. Ces données couvrent une période d'environ 30 000 ans du Paléolithique supérieur au Néolithique final (défini ici par la présence de poterie plutôt que par l'économie agricole de subsistance si non indiquée), proviennent de multiples contextes culturels préhistoriques et proviennent de 54 sites archéologiques dans 14 pays : 1 Aurignacien-associé de Belgique et 1 culturellement inaffecté de Roumanie (35-33 ka), 15 Gravettien-associés d'Espagne, de France, de Belgique, de Tchéquie et d'Italie (31-26 ka), 2 Solutréens- individus associés d'Espagne et de France (23-21 ka), 9 individus associés au Magdalénien de France, d'Allemagne et de Pologne (18-15 ka), 4 individus associés à l'Épigravettien d'Italie (17-13 ka), 2 associés à Federmesser des individus originaires d'Allemagne (14 ka),1, Tableau de données étendu 1 , Données supplémentaires 1.A , Informations supplémentaires, section 1 et Fig. 1 supplémentaire ).

a , Emplacements géographiques des individus nouvellement signalés (symboles remplis avec un contour noir) et des individus représentatifs précédemment publiés (étoiles entourées). Les lignes pointillées délimitent les régions géographiques décrites dans le texte. b , Dates radiocarbone calibrées des individus tracées en a. L' axe y montre la moyenne des datations radiocarbone calibrées en milliers d'années (kyr) (Données supplémentaires 1.A ). La ligne pointillée horizontale marque la limite entre le Pléistocène supérieur et l'Holocène. c , tracé MDS des chasseurs-cueilleurs européens basé sur 1 − f 3(Mbuti ; pop1, pop2). Les dimensions sont calculées à l'aide de groupes ou d'individus de chasseurs-cueilleurs nouvellement signalés et précédemment publiés avec plus de 30 000 SNP. Le regroupement détaillé des individus représentés par des cercles de couleur vides est décrit dans les données supplémentaires 1.I.
(...) Contrairement à la proposition de Fu et al. 4 mais en accord avec Petr et al. 17, nous n'observons pas de diminution substantielle de l'ascendance néandertalienne chez la plupart des chasseurs-cueilleurs européens au fil du temps (Informations supplémentaires, section 6 et Figs. 15 à 17 supplémentaires). Cela fournit un soutien supplémentaire au modèle sans déclin à long terme de l'ascendance néandertalienne à l'échelle du génome chez les humains modernes après leur introgression 18.
Avant la LGM
La culture du Gravettien était l'une des cultures du Paléolithique supérieur les plus répandues dans l'ouest de l'Eurasie avant le LGM 19. Elle est souvent considérée comme une mosaïque culturelle paneuropéenne avec des variations régionales du matériel aux productions symboliques 20, 21. Dans ce cadre débattu, les individus associés au Gravettien ont été suggérés pour représenter une population biologiquement homogène sur la base de données craniométriques et génomiques 4, 22. Cependant, les génomes associés au Gravettien publiés proviennent d'Europe centrale et méridionale, laissant le profil génétique des groupes humains associés au Gravettien d'Europe occidentale et du sud-ouest non décrit.
Pour avoir une vue d'ensemble du contexte génomique des chasseurs-cueilleurs européens avant le LGM, nous avons utilisé la mise à l'échelle multidimensionnelle (MDS) pour tracer une matrice de dissemblance des statistiques f 3 de l'exogroupe par paires sous la forme 1 − f 3 (Mbuti; pop1, pop2) (Fig. 2a). Cette parcelle révèle la présence de trois groupements distincts : (1) un groupe pré-40 ka avec des individus des sites d'Ust'Ishim, Bacho Kiro, Zlatý kůň et Peştera cu Oase, (2) un groupe de Věstonice comprenant des individus associés aux sites Gravettiens d'Europe centrale, orientale et méridionale (Dolní Věstonice, Pavlov, Krems-Wachtberg, Paglicci et Ostuni), et (3) un groupe Fournol (ci-après, groupe Fournol ou ascendance) comprenant des individus associés au Gravettien provenant de sites européens de l'ouest et du sud-ouest (Ormesson , La Rochette, Fournol et deux sites de grottes Serinyà (Mollet III et Reclau Viver)). Le cluster de Věstonice décrit précédemment, comprenant un individu récemment signalé âgé de 29 000 ans de la grotte de Paglicci (Paglicci 12) dans le sud de l'Italie, est étroitement lié aux génomes précédemment publiés de Sunghir et Kostenki 12 dans l'ouest de la Russie, 4, 23. L'amas de Fournol nouvellement défini est étroitement lié aux individus associés à l'Aurignacien de Belgique datés de 35 ka (Goyet Q116-1 et l'individu Goyet Q376-3 nouvellement signalé). Notamment, et contrairement au rapport de Fu et al. 4, une autre population associée au Gravettien d'Europe centrale-occidentale (Goyet en Belgique, n = 6 individus) est à la fois géographiquement et génétiquement intermédiaire entre les amas de Věstonice et de Fournol. La similitude entre Goyet Q116-1 et Goyet Q376-3 et le cluster Fournol est également observée au niveau de l'ADNmt, les deux groupes comprenant des individus porteurs de l'haplogroupe M d'ADNmt, qui n'a pas été trouvé chez les individus européens après le LGM 24 Extended Données Fig.1 et 2).
 a, tracé MDS des individus pré-LGM. Le groupe pré-40 ka et les amas de Fournol et Věstonice sont marqués par des zones ombrées de différentes couleurs. Les individus et les groupes sont tracés avec les mêmes couleurs et symboles que sur la Fig. 1 et les noms sont indiqués à côté des symboles. b, les individus associés au Gravettien forment deux groupes distincts, avec des individus du centre-est et du sud de l'Europe faisant partie de l'amas de Věstonice et des individus de l'ouest et du sud-ouest de l'Europe faisant partie de l'amas de Fournol. Dans le centre-ouest de l'Europe, les individus associés au Gravettien de Goyet montrent une affinité avec les deux groupes. Les barres d'erreur montrent 1 × sem (noir) ou 3 × sem (gris) des valeurs de f 4 estimées à partir d'une analyse jackknife de bloc de 5 cM. c, Modélisation de graphes de mélange des principales lignées de chasseurs-cueilleurs européens pré-LGM créées à l'aide de qpGraph.
a, tracé MDS des individus pré-LGM. Le groupe pré-40 ka et les amas de Fournol et Věstonice sont marqués par des zones ombrées de différentes couleurs. Les individus et les groupes sont tracés avec les mêmes couleurs et symboles que sur la Fig. 1 et les noms sont indiqués à côté des symboles. b, les individus associés au Gravettien forment deux groupes distincts, avec des individus du centre-est et du sud de l'Europe faisant partie de l'amas de Věstonice et des individus de l'ouest et du sud-ouest de l'Europe faisant partie de l'amas de Fournol. Dans le centre-ouest de l'Europe, les individus associés au Gravettien de Goyet montrent une affinité avec les deux groupes. Les barres d'erreur montrent 1 × sem (noir) ou 3 × sem (gris) des valeurs de f 4 estimées à partir d'une analyse jackknife de bloc de 5 cM. c, Modélisation de graphes de mélange des principales lignées de chasseurs-cueilleurs européens pré-LGM créées à l'aide de qpGraph.
(...) Ces résultats montrent que certaines, mais pas toutes, des ascendances génomiques présentes en Europe entre environ 40 ka et 30 ka ont survécu dans les populations associées au Gravettien étudiées jusqu'à présent. L'ascendance Kostenki (et le groupe Sunghir) a contribué au groupe de Věstonice précédemment décrit représenté par des individus associés au Gravettien du centre-est et du sud de l'Europe 4. En revanche, le profil génétique Goyet Q116-1 a donné naissance au cluster Fournol nouvellement décrit identifié chez des individus associés au Gravettien d'Europe de l'Ouest et du Sud-Ouest. Notamment, cette distinction génétique coïncide avec des dissemblances dans la pratique mortuaire parmi les individus associés au Gravettien génétiquement analysés de différentes parties de l'Europe. Les individus de l'ouest et du sud-ouest de l'Europe liés au groupe de Fournol sont systématiquement déposés dans des sites de grottes et présentent parfois des marques anthropiques, tandis que les individus liés au groupe de Věstonice sont enterrés avec des objets funéraires et / ou des ornements personnels et de l'ocre en plein air ou des sites de grottes dans le centre- Europe orientale et méridionale, respectivement (Figures supplémentaires 29 à 32 et Tableau supplémentaire 4). L'individu le plus ancien de l'amas de Fournol est Ormesson 2988 du nord-est de la France (31 ka, Gravettien ancien/moyen), alors qu'un groupe gravettien de Goyet en Belgique (27 ka, Gravettien tardif) se trouve être un mélange entre les amas de Věstonice et de Fournol. Cela suggère qu'entre le Gravettien précoce / moyen et tardif, il y a eu une expansion d'est en ouest de l'ascendance associée à Věstonice qui a atteint le centre-ouest de l'Europe et a créé un cline de mélange longitudinal entre ces deux populations pré-LGM génétiquement distinctes.
LGM dans le sud-ouest et l'ouest de l'Europe
La culture solutréenne est temporellement intermédiaire entre les cultures gravettienne et magdalénienne (ou badegoulienne) et se retrouve dans le sud-ouest et l'ouest de l'Europe, considérés comme des refuges climatiques pour les populations humaines au cours du LGM 25 , 26 . Cependant, la mesure dans laquelle les groupes associés à la culture solutréenne sont en continuité génétique avec les populations antérieures et ultérieures de la même région est inconnue car aucune donnée génomique d'individus associés au solutréen n'a été rapportée auparavant. Les deux génomes nouvellement séquencés d'individus associés au Solutréen (Le Piage II (23 ka) du sud-ouest de la France et La Riera (niveau 14, 21 ka) du nord de l'Espagne) montrent une affinité généralisée avec les membres des groupes Fournol et GoyetQ2 dans l'exogroupe f3 -statistiques (Données supplémentaires 2.A ). Dans la parcelle MDS, l'individu Le Piage II est particulièrement proche des individus appartenant au cluster Fournol, suggérant une continuité génétique locale de cette ascendance au cours du LGM (Fig. 13 supplémentaire ) . Les statistiques de F 4 confirment ce point de vue, révélant que Le Piage II est plus étroitement lié au groupe de Fournol qu'au groupe de Věstonice ( f 4 (Mbuti, Le Piage II; Věstonice, Fournol 85) ≫ 0, Z = 6,58). Nous avons également comparé son affinité avec El Mirón (nord de l'Espagne), le plus ancien individu associé au Magdalénien séquencé à ce jour (19 ka). F-les statistiques suggèrent que Le Piage II est génétiquement intermédiaire entre Fournol 85 et El Mirón (Données supplémentaires 2.D ). De plus, des études antérieures ont montré qu'El Mirón porte une contribution génétique du groupe Villabruna, qui se trouve chez les individus associés à l'épigravettien d'Italie 4, 15. El Mirón a une similitude significativement plus élevée avec le cluster de Villabruna que Fournol 85 et Le Piage II, tandis que l'affinité avec le cluster de Villabruna dans Le Piage II n'est pas significativement plus élevée que dans Fournol 85 (Données supplémentaires 2.D). Dans l'ensemble, l'individu Le Piage II associé au Solutréen relie l'ascendance Fournol précédente à l'ascendance suivante trouvée à El Mirón, fournissant une preuve directe de la continuité génétique dans tout le LGM dans le sud-ouest et l'ouest de l'Europe. Ces régions européennes constituent donc des refuges climatiques où les populations humaines ont survécu pendant le LGM.
Post-LGM dans la péninsule italienne
Après le LGM, la culture épigravettienne s'est répandue dans le sud et le sud-est de l'Europe. Malgré les discussions croissantes sur sa nature 27, 28, la culture épigravettien a été traditionnellement supposée être le résultat d'une transition du Gravettien local précédent 29. Cependant, le niveau de continuité génétique entre les individus associés à ces cultures et la structure de la population parmi les individus associés à l'épigravettien n'ont pas été pleinement explorés. Ici, nous rapportons les données génomiques de 4 individus, dont 3 génomes vieux d'environ 13 000 ans du nord-est de l'Italie (Pradis 1), du nord-ouest de l'Italie (Arene Candide 16) et de la Sicile (San Teodoro 2), ainsi qu'une couverture accrue à l'échelle du génome de Tagliente 2 15 daté de 17 ka.
Dans le diagramme MDS, nous constatons que tous les individus nouvellement et précédemment signalés associés à l'Épigravettien appartiennent au groupe Villabruna 4 (Fig. 1c ). Une série de statistiques de symétrie f 4 confirme que tous les individus associés à l'épigravettien sont cladaux et ne partagent pas d'affinité excessive avec des ancêtres locaux (Paglicci 12) ou non locaux (Goyet Q116-1, Kostenki 14, Mal'ta 1 ou Věstonice) (Données supplémentaires 2.F ). De plus, aucun des individus associés à l'Épigravettien n'a plus d'affinité avec le sud de l'Europe qu'avec les groupes associés au Gravettien d'Europe centrale et orientale, comme le montre f 4 (Mbuti, individu/groupe associé à l'Épigravettien ; Věstonice, Paglicci 12) qui est cohérent avec 0 (Données supplémentaires 2.G ).
Ensuite, nous avons étudié les relations génétiques entre les individus associés à l'épigravettien dans la péninsule italienne, en reconstruisant une phylogénie basée sur une matrice de distances génétiques f 2 par paires (Fig. 3a et Fig. 9 supplémentaire ) et en testant l'affinité relative entre eux à l'aide de f 4 -statistiques sous la forme f 4 (Mbuti, Epigravettien A; Epigravettien B, Epigravettien C) (Données supplémentaires 2.E). La topologie déduite révèle un modèle phylogéographique indépendamment des âges individuels. En particulier, l'individu de 13 ka Pradis 1 du nord-est de l'Italie représente la lignée la plus basale par rapport à tous les autres individus associés à l'épigravettien, y compris les génomes plus anciens de Tagliente 2 et Villabruna du nord de l'Italie (groupe 1). Les individus du nord-ouest de l'Italie (Arene Candide 16), du centre de l'Italie (Continenza) et de la Sicile appartiennent à une branche phylogénétiquement plus dérivée (groupe 2), qui s'est ensuite diversifiée en une branche composée uniquement de chasseurs-cueilleurs siciliens (groupe 3). En Sicile, l'individu Oriente C de 14 ka montre une plus grande affinité avec le groupe Uzzo de 10 ka beaucoup plus jeune mais géographiquement plus proche 30 ( n = 2) qu'avec l'individu presque contemporain de San Teodoro 2 de l'est de la Sicile.
 a, Structure de la population parmi les populations associées à l'épigravettien révélée par un arbre de jonction voisin basé sur les distances génétiques f 2 par paires. Les étiquettes des branches indiquent des longueurs de dérive uniques ; les points noirs font référence aux individus avec des données nouvellement générées et les points blancs font référence aux génomes publiés précédemment ; la position de chaque nœud n'implique pas l'emplacement où la scission a eu lieu. b, Diversité de la population montrée par PMR entre les individus de différents groupes. Le regroupement des populations associées à l'Épigravettien est représenté avec la même couleur dans un. Le losange aux contours noirs matérialisé dans le groupe du Gravettien montre la PMR entre les deux individus associés au Gravettien du sud de l'Italie (Paglicci 12 et Ostuni 1). Dans la boîte à moustaches, la ligne centrale est la médiane, les limites de la boîte délimitent la plage interquartile et les moustaches s'étendent jusqu'aux valeurs maximales et minimales, à l'exclusion des valeurs aberrantes. La taille de l'échantillon des paires individuelles incluses dans chaque groupe est indiquée dans les données supplémentaires 3.A.
a, Structure de la population parmi les populations associées à l'épigravettien révélée par un arbre de jonction voisin basé sur les distances génétiques f 2 par paires. Les étiquettes des branches indiquent des longueurs de dérive uniques ; les points noirs font référence aux individus avec des données nouvellement générées et les points blancs font référence aux génomes publiés précédemment ; la position de chaque nœud n'implique pas l'emplacement où la scission a eu lieu. b, Diversité de la population montrée par PMR entre les individus de différents groupes. Le regroupement des populations associées à l'Épigravettien est représenté avec la même couleur dans un. Le losange aux contours noirs matérialisé dans le groupe du Gravettien montre la PMR entre les deux individus associés au Gravettien du sud de l'Italie (Paglicci 12 et Ostuni 1). Dans la boîte à moustaches, la ligne centrale est la médiane, les limites de la boîte délimitent la plage interquartile et les moustaches s'étendent jusqu'aux valeurs maximales et minimales, à l'exclusion des valeurs aberrantes. La taille de l'échantillon des paires individuelles incluses dans chaque groupe est indiquée dans les données supplémentaires 3.A.
(...) Pour résumer, nos résultats mettent en évidence un renouvellement génétique dans la péninsule italienne de l'amas de Věstonice associé au Gravettien par l'amas de Villabruna associé à l'Épigravettien qui pourrait être corrélé aux discontinuités observées dans les archives archéologiques 31. Nous montrons que tous les individus analysés associés à l'Epigravettien portent une ascendance Villabruna homogène, la structure génétique intra-groupe étant principalement déterminée par leur distribution géographique et non temporelle. La reconstruction phylogénétique des génomes associés à l'épigravettien, avec Pradis 1 divergeant plus profondément que tous les autres, indique que le renouvellement a eu lieu bien avant 17 ka - la date du génome Tagliente 2 plus dérivé. Ceci, ainsi que la preuve de l'ascendance Villabruna à El Mirón 19 ka, suggère en outre que cette discontinuité génétique pourrait être le résultat de transformations paléogéographiques et paléoécologiques liées au LGM 32, plutôt qu'à la période de réchauffement Bølling–Allerød 4 , 15 (14,7–12,9 ka). De plus, notre analyse phylogéographique indique que le nord-est de l'Italie est le point d'entrée possible du pool génétique associé à l'épigravettien dans la péninsule italienne. Cette découverte, en conjonction avec l'affinité génétique du groupe de Villabruna avec les ancêtres anciens et actuels du Proche-Orient 4 , 15 , 16 (Informations supplémentaires, section 8 , Fig. 18 supplémentaire et Données supplémentaires 2.O), suggère les Balkans comme source de la population entrante associée à l'Épigravettien. Le LGM aurait ainsi pu créer un corridor au sud des Alpes pour les mouvements humains d'est en ouest qui reliaient génétiquement les populations de chasseurs-cueilleurs des Balkans à la péninsule ibérique, peut-être aussi via des dispersions le long des côtes existantes au niveau de la mer 32.
Post-LGM en Europe occidentale et centrale
La culture magdalénienne a été largement diffusée dans le sud-ouest, l'ouest et le centre de l'Europe après le LGM 33. Malgré cette large répartition géographique, il n'est pas clair si les différents groupes associés à cette culture provenaient d'une population source commune et comment ces groupes étaient génétiquement liés les uns aux autres. Des études antérieures ont identifié deux compositions génétiques différentes chez les individus associés au Magdalénien - le cluster GoyetQ2 comprenant des génomes d'Europe centrale et occidentale datés d'environ 15 ka (de France, de Belgique et d'Allemagne) et l'ascendance de l'individu El Mirón d'Espagne 4 , 14 d'environ 19 ka. Ces deux ancêtres portent une composante génétique éloignée de l'individu Goyet Q116-1 daté de 35 ka, l'individu ibérique montrant également une affinité avec le groupe Villabruna 4 , 14. En co-analysant les données précédemment publiées avec nos génomes nouvellement rapportés associés au Magdalénien de La Marche (18 ka) et Pincevent (15 ka) dans l'ouest et le nord de la France, respectivement, et Maszycka (18-16 ka) dans le sud de la Pologne, nous confirment que l'ascendance Goyet Q116-1 a survécu dans tous les génomes associés au Magdalénien étudiés, à l'exception des individus associés au Gravettien et au Solutréen du sud-ouest et de l'ouest de l'Europe (Fig. 1). Notamment, l'ascendance Fournol fournit un meilleur proxy que Goyet Q116-1 pour la composante génétique trouvée dans le cluster GoyetQ2 et à El Mirón (Données supplémentaires 2.H). Cependant, en utilisant les statistiques f 4, nous montrons que tous les individus associés au Magdalénien, et pas seulement El Mirón, portent une ascendance liée à Villabruna par rapport au groupe Fournol (Données supplémentaires 2.H). Cette affinité est encore plus forte envers les individus associés à l'Épigravettien de l'ouest et du centre de l'Italie et de la Sicile (groupe 2 et groupe 3, respectivement) qu'avec ceux du nord de l'Italie (groupe 1) (Données supplémentaires 2.F).
Nous avons ainsi modélisé les individus appartenant au cluster GoyetQ2 et El Mirón comme un mélange entre les génomes Fournol 85 et Arène Candide 16 comme proxies pour représenter les ascendances Fournol et Villabruna, respectivement, dans les groupes associés au Magdalénien (Fig. 4a). Outre El Mirón, qui a environ 43% d'ascendance Villabruna, tous les autres individus associés au Magdalénien ont une proportion plus faible de cette composante (19-29%) et peuvent donc être attribués au cluster GoyetQ2 (Fig. 4a et Données supplémentaires 3.C ). Ceci est encore validé par f 4 -statistiques de la forme f 4 (Mbuti, Arène Candide 16 ; Goyet Q-2, Individus associés au Magdalénien), qui n'est significativement positif que pour El Mirón, alors que tous les autres individus testés et Goyet Q-2 sont apparentés symétriquement par rapport à Arène Candide 16 (Fig. 26 et données supplémentaires 2.H).
 a, b, Les ascendances des individus du cluster GoyetQ-2 et des chasseurs-cueilleurs (HG) ibériques ( a ) et des individus du cluster Oberkassel ( b ) ont été modélisées à l'aide de qpAdm, avec Fournol 85 et Arene Candide 16 représentant les Fournol et Ascendances Villabruna, respectivement. La longueur de la barre de couleur indique la proportion de chaque ascendance. La barre d'erreur montre la sem des estimations de l'analyse jackknife de blocs de 5 cM. Les détails de la modélisation sont fournis dans les données supplémentaires 3.C.
a, b, Les ascendances des individus du cluster GoyetQ-2 et des chasseurs-cueilleurs (HG) ibériques ( a ) et des individus du cluster Oberkassel ( b ) ont été modélisées à l'aide de qpAdm, avec Fournol 85 et Arene Candide 16 représentant les Fournol et Ascendances Villabruna, respectivement. La longueur de la barre de couleur indique la proportion de chaque ascendance. La barre d'erreur montre la sem des estimations de l'analyse jackknife de blocs de 5 cM. Les détails de la modélisation sont fournis dans les données supplémentaires 3.C.
Nos analyses démontrent que le cluster Fournol est une meilleure source de génomes associés au Magdalénien que Goyet Q116-1. Par conséquent, la plupart des ancêtres trouvés chez ces individus post-LGM remontent probablement à des groupes associés au Gravettien d'Europe de l'Ouest et du Sud-Ouest. L'affinité génétique avec l'ascendance Villabruna est présente chez El Mirón et chez les individus associés au Magdalénien d'Europe occidentale et centrale. Cela suggère que les liens génétiques entre les chasseurs-cueilleurs du sud et du sud-ouest de l'Europe à l'époque du LGM s'étendaient au nord des Pyrénées. Le cluster GoyetQ2 résultant comprend des individus allant de l'ouest de la France à la Pologne dans la période comprise entre 18 et 15 ka. Par conséquent, contrairement aux suggestions précédentes 34, cela démontre que la diffusion post-LGM du Magdalénien était bien associée à des expansions de population vers le nord et le nord-est depuis l'Europe occidentale 35.
Post-14 ka au Néolithique
Des études antérieures ont montré que deux principales ascendances de chasseurs-cueilleurs prédominaient dans la plupart des régions d'Europe après environ 14 ka, à savoir l'ascendance des chasseurs-cueilleurs de l'ouest (WHG), liée au groupe Villabruna, et l'ascendance des chasseurs-cueilleurs de l'est (EHG ) ascendance, montrant une affinité à la fois avec l'ascendance Villabruna et l'ancienne ascendance nord-eurasienne (ANE) trouvée chez les individus sibériens du Paléolithique supérieur 4, 36. Des chasseurs-cueilleurs porteurs d'un profil génétique mixte WHG/EHG ont été séquencés dans diverses régions d'Europe du Nord et de l'Est, ce qui soulève la question de savoir comment ces deux types d'ascendances se sont formées et ont interagi dans le temps et dans l'espace 37, 38, 39, 40.
Dans le diagramme MDS (Fig. 1c ) et une analyse en composantes principales (PCA) de l'ouest de l'Eurasie (Extended Data Fig. 6 et Supplementary Fig. 14), la plupart des individus post-14 ka d'Europe occidentale et centrale se situent à proximité du cluster WHG et ceux d'Europe de l'Est proches du cluster EHG, tandis que l'individu Tutkaul 1 d'Asie centrale se situe à proximité du groupe lié à l'ANE. Les deux individus Oberkassel de 14 ka marquent la présence la plus ancienne d'ascendance WHG au nord des Alpes, que nous renommons donc le groupe Oberkassel (ci-après, groupe ou ascendance Oberkassel), en utilisant le nom du plus ancien individu signalé à ce jour portant une telle ascendance avec plus de couverture unique, pour plus de cohérence 4. Sur la base de f 4-statistiques, nous constatons que les individus affectés au cluster d'Oberkassel sont plus proches du génome d'Arene Candide 16 que tout autre groupe associé à l'épigravettien d'Italie (données supplémentaires 2.F). De plus, le cluster Oberkassel porte à la fois l'ascendance Villabruna et une contribution de l'ascendance GoyetQ2 (Données supplémentaires 2.J). Cela a été confirmé avec qpAdm, dans lequel nous avons pu modéliser tous les individus du cluster d'Oberkassel comme un mélange globalement constant d'environ 75 % d'Arène Candide 16 et 25 % de Goyet Q-2 (ou 90 % d'Arène Candide 16 et 10 % de Fournol 85) ( Fig. 4b et données supplémentaires 3.C). L'observation que les individus post-14 ka d'Europe occidentale et centrale et aussi de Grande-Bretagne 41 portent une composition génétique homogène au lieu d'afficher des mélanges locaux répétés avec l'ascendance GoyetQ2 implique que le profil d'ascendance Oberkassel était déjà largement formé avant sa dispersion. Cela contraste fortement avec l'histoire génétique des chasseurs-cueilleurs ibériques, où la propagation de l'ascendance Villabruna / Oberkassel impliquait de multiples événements de mélange locaux avec des groupes portant des proportions élevées d'ascendance GoyetQ2 14 (Fig. 4 et données supplémentaires 3.C). La continuité génétique de longue durée dans la péninsule ibérique se reflète également dans la préservation jusqu'au Mésolithique de l'haplogroupe C du chromosome Y, qui était prédominant dans les groupes pré-LGM mais rarement trouvé après le LGM dans d'autres parties de l'Europe (Extended Data Figs. 1 et 2).
(...) Nous avons ensuite tenté de modéliser 250 chasseurs-cueilleurs publiés et nouvellement signalés datés de 14 à 5 ka en utilisant qpAdm comme un mélange d'ascendances Oberkassel, Sidelkino, GoyetQ2 et une ascendance maximisée chez les agriculteurs néolithiques anatoliens (ANF), comme une partie considérable de la les génomes séquencés des chasseurs-cueilleurs datent d'environ 8 ka, lorsque l'ascendance ANF a commencé à se répandre à travers l'Europe. Nos résultats montrent que la zone de contact et les schémas de mélange entre les ancêtres Oberkassel et Sidelkino ont changé au fil du temps (Fig. 5). Entre 14 et 8 ka, tous les chasseurs-cueilleurs d'Europe occidentale et centrale ne portaient que l'ascendance Oberkassel, sans contribution détectable du groupe Sidelkino. Plus au nord et à l'est, des individus des pays baltes (Baltic HG), de Scandinavie (SHG), des Balkans (Iron Gates HG) et d'Ukraine (Ukraine HG) portaient déjà une ascendance mixte Oberkassel/Sidelkino 38, 40 avant 8 ka. En outre, ces groupes ont également une affinité avec l'ANF suggérant des processus génétiques plus complexes derrière leur histoire démographique 16. De plus, deux des plus anciens groupes publiés de l'ouest de la Russie appartenant au groupe Sidelkino - Peschanitsa (13 ka) 43et les individus Minino nouvellement signalés (11 ka) - ont montré une affinité supplémentaire avec le groupe d'Oberkassel, peut-être en raison de la variabilité de cette proportion d'ascendance pendant la phase de formation initiale du profil d'ascendance Sidelkino. À l'aide du logiciel DATES, nous avons estimé le mélange entre les ancêtres Villabruna / Oberkassel et ANE chez ces anciens individus liés au groupe Sidelkino à environ 15 à 13 ka (données étendues Fig. 7 et tableau supplémentaire 3), ce qui coïncide à peu près avec la première apparition de l'ascendance Oberkassel en Europe centrale. Cela soulève la possibilité que le remplacement par l'amas d'Oberkassel et la formation de l'amas de Sidelkino aient pu être le résultat d'expansions démographiques influencées par le réchauffement brutal de l'interstade Bølling-Allerød 4 , 24.

Fig 5, a, L'ascendance génétique des chasseurs-cueilleurs datée entre 14 ka et 5,2 ka modélisée à l'aide de qpAdm, avec Oberkassel, Yuzhniy Oleniy Ostrov, Goyet Q-2 et les agriculteurs néolithiques de la Turquie actuelle (sites de Barcın, Menteşe et Boncuklu) représentant Oberkassel ( WHG) (bleu), Sidelkino (EHG) (rouge), GoyetQ2 (orange) et ancêtres fermiers néolithiques anatoliens (vert), respectivement. La date moyenne calibrée est indiquée, avec des diagrammes circulaires indiquant la proportion estimée d'ascendance pour chaque groupe ou individu. Les détails de la modélisation sont fournis dans les données supplémentaires 3.E,F.
L'expansion de l'agriculture de 9, 8, 7,5 et 7 ka est représentée par des nuances vertes Adapté de https://doi.org/10.5281/zenodo.5903165 (CC BY 4.0). b, Fréquences alléliques de différents groupes de chasseurs-cueilleurs (points colorés) sur quatre SNP liés à la couleur de la peau ( SLC24A5 et SLC45A2 ), la couleur des yeux ( HERC2 / OCA2 ) et la persistance de la lactase (LCT). Les points sont des estimations de vraisemblance maximale et les barres d'erreur montrent des intervalles de confiance à 95 % des fréquences alléliques dérivées ( n , le nombre d'individus dans chaque groupe, est fourni dans les données supplémentaires 3.G). Les lignes pointillées montrent les fréquences estimées pour les populations actuelles indiquées du projet 1000 Genomes (CEU, résidents de l'Utah d'ascendance européenne du nord et de l'ouest ; GBR, britannique ; IBS, espagnol ; TSI, toscan) 37. Des détails sur les estimations de fréquence d'allèles sont fournis dans Informations supplémentaires, section 12, Fig. 27 et 28 et données supplémentaires 3.G.
À partir d'environ 8 ka, nous commençons à observer des événements de mélange avec l'ascendance Sidelkino en Europe centrale. Ceci est d'abord détecté chez un individu de Gross Fredenwalde dans le nord-est de l'Allemagne et atteint environ 10% chez la plupart des individus de chasseurs-cueilleurs européens par la suite (Extended Data Fig. 8 ). Peu après 8 ka, l'ascendance Sidelkino était absente de l'est de l'Espagne, mais elle avait déjà atteint le nord de la péninsule ibérique parallèlement à une augmentation de l'ascendance Oberkassel (Fig. 5). À l'inverse, une ascendance Oberkassel supplémentaire est identifiée en Europe de l'Est par au moins 7,5 ka dans les génomes nouvellement générés de Minino I et Yazykovo de la région de la haute Volga, alors qu'un individu de 1000 ans de Minino I n'avait pas cette composante génétique. Considérant un signal de réservoir d'eau douce dans la région de la haute Volga faisant apparaître des datations au radiocarbone sur des restes humains jusqu'à environ 500 ans de plus que leur âge réel 44, il pourrait y avoir un intervalle de plus de 1000 ans entre la première preuve de mélange chez les chasseurs-cueilleurs d'Europe centrale d'ascendance Sidelkino et les chasseurs-cueilleurs d'Europe de l'Est d'ascendance Oberkassel. Cependant, des génomes supplémentaires intermédiaires dans le temps et dans l'espace sont nécessaires pour évaluer si ces deux événements de mélange étaient indépendants ou faisaient partie d'un processus démographique commun.
Après 7,5 ka, alors que l'ascendance ANF avait atteint les régions au nord des Alpes, les individus porteurs d'un profil génétique de chasseurs-cueilleurs étaient principalement limités à la frange nord de l'Europe (Fig. 5 ) . Au cours de cette période, le mélange d'ascendance Oberkassel s'est répandu plus à l'est, atteignant Samara il y a environ 6,5 ka, et une augmentation de l'ascendance Sidelkino a été détectée chez les chasseurs-cueilleurs de la région de la Baltique, qui était auparavant associée à la transition de la culture Narva à la culture Peigne en Céramique 38, 39 (Extended Data Fig. 8). En Europe centrale, le mélange avec l'ascendance ANF est devenu très courant mais pas omniprésent, indiquant la coexistence de sociétés de chasseurs-cueilleurs et d'agriculteurs sans mélange pendant plusieurs centaines d'années. Le plus jeune individu porteur d'une grande partie de l'ascendance des chasseurs-cueilleurs dans l'ensemble de données analysé provient d'Ostorf, dans le nord de l'Allemagne, daté d'environ 5,2 ka (> 90 % du groupe Oberkassel plus les composants du groupe Sidelkino) (Données supplémentaires 3.F). Les individus de ce site pourraient marquer l'une des dernières occurrences d'ascendances aussi élevées liées aux chasseurs-cueilleurs, quelques siècles seulement avant l'émergence de l'âge du bronze européen.
Sur la base des statistiques de l'ACP et de l'exogroupe f 3, l'individu néolithique Tutkaul 1 du Tadjikistan est étroitement lié aux individus du Paléolithique supérieur du centre-sud de la Sibérie (Afontova Gora 3 (AG3) et Mal'ta 1), et à peu près contemporains des chasseurs-cueilleurs (Tyumen et Sosnoviy) de la Sibérie occidentale, tous deux porteurs de fortes proportions d'ascendance ANE 45 (Fig. 1c et Extended Data Fig. 6). Nous avons testé l'affinité de Tutkaul 1 avec les populations anciennes et modernes du monde entier par rapport à AG3. Contrairement aux chasseurs-cueilleurs de Sibérie occidentale, Tutkaul 1 ne porte pas d'ascendance eurasienne extra-orientale, mais montre une affinité avec les agriculteurs néolithiques iraniens et certaines populations plus jeunes d'Iran et de la région de Turan (Données supplémentaires 2.L). À l'inverse, les individus du groupe Sidelkino sont génétiquement plus proches d'AG3 que de Tutkaul 1. Cela suggère que l'individu néolithique nouvellement signalé d'Asie centrale porte une ascendance qui pourrait être un bon indicateur de la contribution liée à l'ANE à l'Iran et à la région de Turan, il y a environ 5,5 ka mais pas aux chasseurs-cueilleurs à peu près contemporains d'Europe de l'Est.
En résumé, nous décrivons la formation et l'interaction entre les amas d'Oberkassel et de Sidelkino, les deux principales ascendances de chasseurs-cueilleurs présentes en Europe à partir de 14 ka. La similitude génomique du groupe d'Oberkassel avec Arene Candide 16 dans le nord-ouest de l'Italie pourrait impliquer que l'ascendance associée à l'épigravettien s'est propagée du sud à l'Europe centrale en passant par le côté ouest de la région alpine. L'ascendance Sidelkino a également émergé vers 14 ka avec sa première preuve directe en Europe de l'Est 43 daté de 13 ka. Le niveau croissant de mélange entre les populations distinctes de chasseurs-cueilleurs à partir d'environ 8 ka indique une mobilité intensifiée de ces groupes de butineurs. Cela pourrait avoir été en partie déclenché par l'expansion concomitante des agriculteurs néolithiques à travers l'Europe et/ou par des facteurs environnementaux, tels que l'événement climatique vers 8,2 ka, le plus grand refroidissement brutal de l'hémisphère nord au cours de l'époque holocène 46, 47.
Variantes phénotypiquement pertinentes
En tirant parti de la taille considérablement accrue de l'échantillon, nous avons étudié des groupes de chasseurs-cueilleurs génétiquement distincts pour les fréquences d'allèles à des locus sélectionnés qui sont connus pour être associés à des traits phénotypiques spécifiques chez les Européens actuels (Fig. 5b et Fig. 27 et 28 supplémentaires). Conformément aux découvertes précédentes, aucun des groupes analysés ne montre l'allèle dérivé au SNP rs4988235 sur le gène LCT, qui est responsable de la persistance de la lactase. Comme indiqué précédemment 37, nous trouvons une grande variation de fréquence dans les allèles liés à la pigmentation de la peau et des yeux parmi les groupes de chasseurs-cueilleurs post-LGM. Pour le SNP associé à la couleur claire des yeux ( HERC2/OCA2(rs12913832)), les individus des groupes Villabruna, Oberkassel, Baltic HG et SHG présentent des fréquences élevées de l'allèle dérivé (>90%), qui est responsable du phénotype œil vert ou bleu, alors que le groupe Sidelkino, Ukraine HG et Les groupes Iron Gates HG présentent une faible occurrence de cet allèle (10 à 25%). Au lieu de cela, pour les deux SNP associés à la couleur de la peau ( SLC24A5 (rs1426654) et SLC45A2 (rs16891982)), le cluster Sidelkino et les groupes Ukraine HG montrent une fréquence plus élevée (> 90 % pour SLC24A5 et 29 à 61 % pour SLC45A2) des allèles dérivés liés à la couleur claire de la peau, par rapport aux clusters Oberkassel et Villabruna, où ces allèles sont presque totalement absents (<1%). Sur la base de la variation génétique des Européens actuels, cela pourrait impliquer des différences phénotypiques entre les populations de chasseurs-cueilleurs post-14 ka à travers l'Europe, les individus du groupe Oberkassel (occident) présentant peut-être une peau plus foncée et des yeux plus clairs, et les individus du groupe Sidelkino (Est) peut-être une peau plus claire et des yeux plus foncés.
Nous fournissons cinq nouvelles informations sur l'histoire génomique des populations de chasseurs-cueilleurs sur une période de 30 000 ans, du Paléolithique supérieur au Néolithique.
Premièrement, nous montrons que les individus associés à la culture gravettienne à travers l'Europe ne constituaient pas une population biologiquement homogène. Culturellement, cependant, nous voyons à la fois des tendances générales répandues, telles que l'armement et certains arts mobiliers 48, et d'autres aspects qui ont un caractère plus régional, tels que les pratiques mortuaires (Informations complémentaires, section 13), diverses originalités dans les matériaux lithiques et organiques durs. trousses à outils et ornements 20, 21. L'ascendance trouvée chez les individus associés à la culture aurignacienne précédente d'Europe centrale (ascendance GoyetQ116-1) a donné naissance à des individus associés au Gravettien d'Europe de l'Ouest et du Sud-Ouest. Cette ascendance dérivée - l'amas Fournol - a survécu au LGM chez des individus associés au Solutréen, peut-être dans le refuge climatique franco-cantabrique 25, conduisant à des populations ultérieures associées à la culture magdalénienne (amas GoyetQ2 et El Mirón). À l'inverse, l'ascendance trouvée chez les individus d'Europe de l'Est d'avant 30 ka (groupe de Kostenki et groupe de Sunghir) a contribué aux individus associés au Gravettien d'Europe centrale et méridionale (groupe de Věstonice), ces derniers sans descendants récupérés dans les populations post-LGM de ces régions.
Deuxièmement, l'ascendance des individus associés à la culture Epigravettien (cluster de Villabruna), dont on a découvert qu'ils connectaient génétiquement les chasseurs-cueilleurs d'Europe et du Proche-Orient, a atteint le sud de l'Europe bien avant la transition entre l'Epigravettien précoce et tardif 4, 15 et peut-être aussi tôt comme la transition Gravettien-Epigravettien. Une reconstruction phylogéographique de différentes lignées portant cette ascendance suggère en outre son point d'entrée dans le nord-est de l'Italie depuis les Balkans, suivi d'une expansion nord-sud dans la péninsule italienne parallèlement à un déclin de la population par des goulots d'étranglement séquentiels.
Troisièmement, les individus associés au Magdalénien non seulement de la péninsule ibérique mais aussi du reste de l'Europe portent une ascendance associée à l'épigravettien (groupe de Villabruna). Les analyses génétiques d'individus d'Europe occidentale associés à la culture badegoulienne précédente pourraient fournir des indices sur les processus qui ont conduit à la formation du cluster GoyetQ2. Comme l'ont déduit les archives archéologiques 35, la propagation du Magdalénien à travers l'Europe est liée aux expansions de population post-LGM du sud-ouest vers le nord et le nord-est et non aux mouvements depuis les refuges du sud-est 34.
Quatrièmement, nous étendons la découverte d'un renouvellement génétique à grande échelle dès 14 ka chez les chasseurs-cueilleurs d'Europe centrale et occidentale associés à de multiples techno-complexes - Federmesser, Azilian et d'autres groupes du Paléolithique final 4 — malgré une grande continuité technologique avec le Magdalénien supérieur précédent. Cette ascendance largement distribuée (le groupe d'Oberkassel (également connu sous le nom de WHG)) est plus étroitement liée à un individu associé à l'épigravettien du nord-ouest de l'Italie, ce qui suggère que son expansion en Europe continentale pourrait avoir commencé à partir de l'ouest - et non de l'est - du côté des Alpes. De plus, le remplacement génétique presque complet du pool génétique associé au Magdalénien soulève l'hypothèse que certaines parties de l'Europe étaient peuplées de manière différentielle lors de la variation climatique abrupte commençant vers 14,7 ka avec la période de réchauffement de Bølling – Allerød , créant des zones où les populations du sud de l'Europe pourraient se développer.
Cinquièmement, l'ascendance Oberkassel en Europe occidentale et centrale et l'ascendance Sidelkino en Europe orientale sont restées largement isolées pendant près de 6 000 ans jusqu'à ce que des interactions génétiques soient observées pour la première fois - vers environ 8 ka dans le nord-est de l'Allemagne, éventuellement associées à des échanges culturels le long de la Baltique 49 et aux environs de 7,5 ka dans la région de la haute Volga, peut-être lié à la diffusion de la poterie dans la région 50.
En conclusion, notre étude révèle que l'ouest et le sud-ouest de l'Europe ont servi de refuge climatique à la persistance des groupes humains lors de la phase la plus froide de la dernière période glaciaire alors que les populations de la péninsule italienne et de la plaine d'Europe orientale ont été génétiquement bouleversées, remettant en cause le rôle de ces populations et régions comme refuges glaciaires pour les humains. L'ascendance entrante de Villabruna est devenue plus tard l'ascendance de chasseurs-cueilleurs la plus répandue à travers l'Europe. D'autres études paléogénomiques sur les individus du Paléolithique supérieur des Balkans seront essentielles pour comprendre si le sud-est de l'Europe représente la source de l'ascendance Villabruna et un refuge climatique pour les populations humaines pendant le LGM.
La suite de l'article scientifique décrit les méthodes utilisées pour les datations, échantillonages et séquences d'ADN et autres. D'autres données sont ici disponibles pour les chercheurs : https://www.nature.com/articles/s41586-023-05726-0#MOESM2
Un autre article va suivre concernant une autre étude sur les mêmes périodes, mais concernant la comparaison des industries lithiques des chasseurs-cueilleurs.
Yves Herbo, Traductions et compilations de données, Sciences-Faits-histoires, 19-03-2023


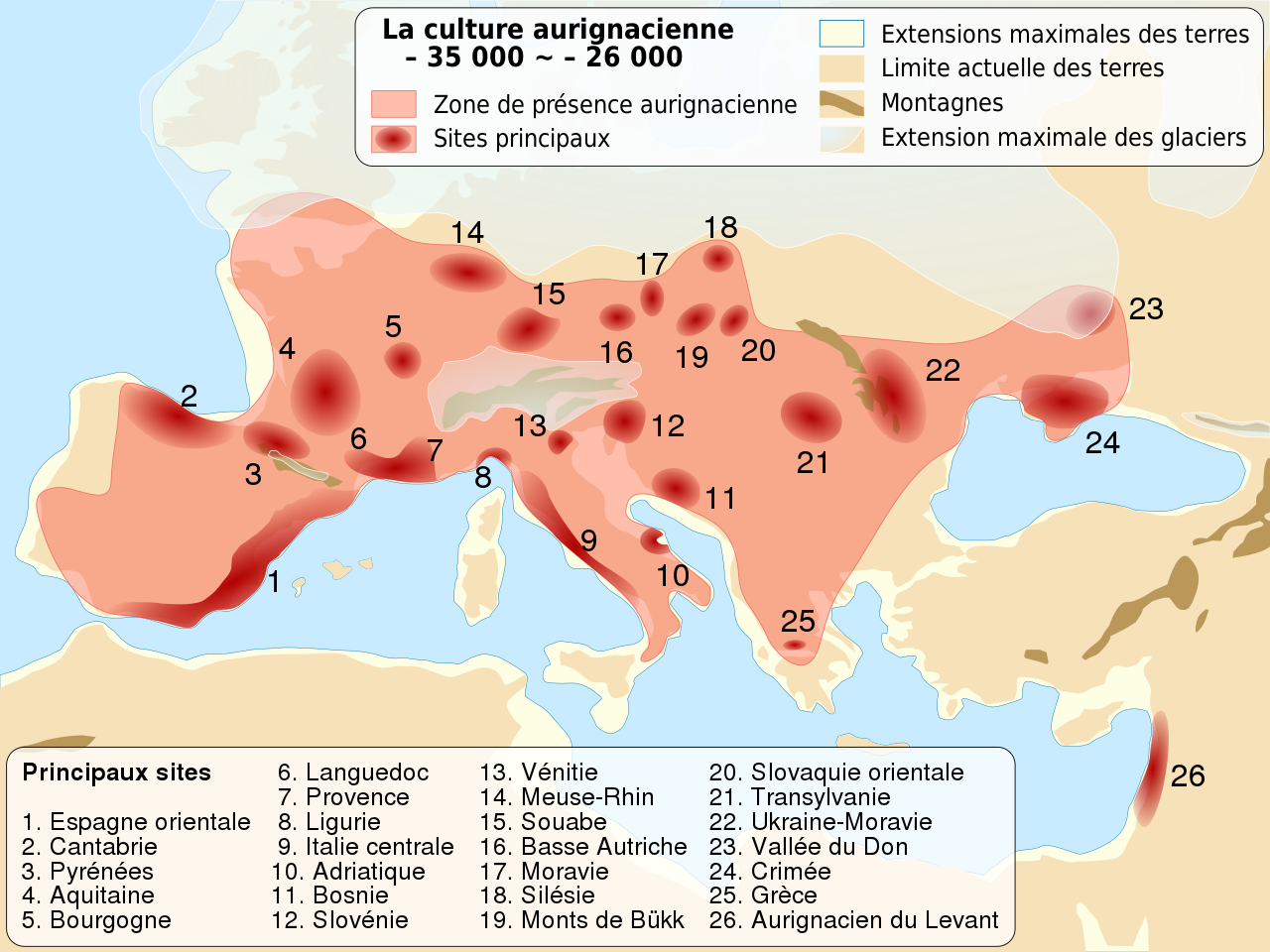 Carte de localisation de la culture Homo Sapiens de l'Aurignacien, il y a entre 37 000 et 28 000 ans. © Sémhur / Wikimedia Commons - CC BY-SA 4.0
Carte de localisation de la culture Homo Sapiens de l'Aurignacien, il y a entre 37 000 et 28 000 ans. © Sémhur / Wikimedia Commons - CC BY-SA 4.0








